Art: Gesamtheit aller Lebewesen, die untereinander bei Verschiedengeschlechtlichkeit und Gesundheit unbegrenzt fortpflanzungsfähig sind (diese entscheidende Voraussetzung heißt »Panmixie«).
Einschränkungen dieser Fortpflanzungsfähigkeit zwischen verschiedengeschlechtlichen Individuen können an vielen Stellen einsetzen:
-
sie paaren sich nicht (was wieder unterschiedliche Gründe haben kann: geographische Trennung; zeitliche Trennung [z.B. durch unterschiedliche Schlupfzeiten bei kurzer Lebensdauer]; ökologische Trennung an den zur Paarung aufgesuchten Stellen, z.B. Laichplätzen; unterschiedliche sexuelle Auslöser, z.B. Zeichnungselemente, Lautäußerungen, Lichtsignale oder Verhaltenssequenzen);
-
sie paaren sich zwar, erzeugen aber keine oder keine fortpflanzungsfähigen Nachkommen (z.B. weil diese steril bleiben, als Embryonen sterben oder das  bei der Paarung stirbt);
bei der Paarung stirbt);
-
sie paaren sich und erzeugen auch fortpflanzungsfähige Nachkommen, welche aber dauerhaft geminderte Fortpflanzungschancen haben und daher im Laufe der Generationen keine weiteren Nachkommen hinterlassen; dies bewirken körperliche Eigenschaften oder Verhaltensweisen, die die Wirksamkeit sexueller Auslöser beeinträchtigen oder das Überleben bis zur Paarung (evtl. auch Brutpflege) weniger wahrscheinlich machen.–
Zwar pflanzen sich viele Lebewesen nicht sexuell fort, entweder primär wie die Mehrheit aller einzelligen Lebewesen (aus einer einzigen, sicherlich schon mit Sexualität versehenen Population von ihnen sind alle Vielzeller hervorgegangen) oder sekundär wie viele Pflanzen, die sich entweder nur durch Knospung oder nur durch Selbstbestäubung vermehren, daneben etliche »Würmer«, Insekten, Schnecken, Fische und Eidechsen – der »Ausstieg« aus der Sexualität kann an den verschiedensten Stellen des Natürlichen Systems (s.u.) erfolgen, hat aber niemals besonders erfolgreiche Taxa (s.u.) hervorgebracht. Alle Lebewesen, die nicht auf sexuellem Wege entstanden sind, heißen Klone; sie sind getreue genetische Kopien ihrer Ausgangsindividuen, sozusagen »jüngere eineiige Zwillinge«. Da allerdings bei diesem Vorgang genausogut Kopierfehler auftreten können wie bei den entsprechenden Vorgängen im Zusammenhang mit der sexuellen Fortpflanzung, kann auch bei den sich klonierend fortpflanzenden Lebewesen eine  Evolution eintreten, sie verläuft nur prinzipiell langsamer (was die einzelligen, besonders die prokaryotischen Lebewesen allerdings durch ihre ungeheure Zahl und Vermehrungsgeschwindigkeit gegenüber den komplizierter gebauten ausgleichen).
Evolution eintreten, sie verläuft nur prinzipiell langsamer (was die einzelligen, besonders die prokaryotischen Lebewesen allerdings durch ihre ungeheure Zahl und Vermehrungsgeschwindigkeit gegenüber den komplizierter gebauten ausgleichen).
Diese Tatsache, daß nicht alle Lebewesen einer Art (im strengen Sinne der obigen Definition) angehören (die einzige seltene und sekundäre sonstige Ausnahme neben dem Klon ist das Klepton), hat einige rückwärtsgewandte, letztlich der Religion verpflichtete Wortführer zu spitzfindigen Plädoyers zugunsten einer Rückkehr zum vorwissenschaftlichen subjektiven Artbegriff veranlaßt – etwa »Art ist, was dem Beobachter ähnlich erscheint«, womit jeder wissenschaftlichen Biologie ein Ende bereitet wäre. Dies zeigt schon die einfachste Überlegung: erstens gibt es – die oben gegebene wissenschaftliche Begriffsbestimmung stets vorausgesetzt – zahlreiche »kryptische Arten«, d.h. Arten, deren Getrenntheit keinem Betrachter anhand ihrer Erscheinung auffallen kann (weil ihre Paarungsbarrieren kein morphologisches Substrat besitzen, sondern anders, z.B. durch Geruch oder Lautäußerungen bewirkt werden – bekanntestes Beispiel für letzteres sind Fitis [Phylloscopus trochilus] und Zilpzalp [Ph. collybita] [zu den wissenschaftlichen Benennungsprinzipien s.u.]; manchmal unterscheiden sie sich morphologisch auch nur in ihren Larvenformen oder gewissen Stadien davon, die lange Zeit unbeachtet bleiben können – berühmtestes Beispiel davon sind die Tagfalter Goldene Acht [Colias hyale] und ihr etwas südlicheres Pendant [C. australis]); andererseits gibt es viele Arten, deren äußeres Erscheinungsbild so variabel ist – wobei nicht einmal Übergangsformen existieren müssen –, daß kein Betrachter ohne Verhaltensbeobachtung und Zuchtexperiment auf den Gedanken käme, es mit Individuen der gleichen Art zu tun zu haben (als Beispiel genügt unser häufiger, keineswegs immer zweipunktiger Zweipunktiger Marienkäfer; US-Amerikaner mögen an ihre Hummelart Bombus rufocinctus denken).
Variabilität und Gleichförmigkeit der Eigenschaften der Mitglieder einer Art, also auch ihre morphologischen Eigenschaften, sind durch die Gene ihres Genpools gegeben; wenn diese nicht sexuell ausgetauscht werden, genügt als Kriterium für die Analogie zur Art also die Übereinstimmung des Genbestandes, um die Analogie zur Artzugehörigkeit festzuschreiben. Schließlich haben sich die Arten im strengen Sinne des Wortes ja nur durch Genaustausch unter zuvor ausschließlich durch Klonierung vermehrten Individuen entwickelt, was deren weitgehend übereinstimmenden Bestand und dessen kompatible Anordnung zur Voraussetzung hatte; es reicht daher für den praktischen Bedarf ebenso wie für das theoretische Verständnis völlig aus, die entsprechenden nur oder vorwiegend durch Klonierung vermehrten Gesamtheiten weiterhin als »Arten« zu bezeichnen, im Bedarfsfall als »Art-Analoga«.
Die Feststellung, ob mehrere Lebewesen oder Gruppen von Lebewesen verschiedenen Arten angehören oder nicht, ist somit eine empirische Frage und nicht eine der Konvention. Sie kann richtig oder falsch beantwortet werden; oft ist die Beantwortung sehr schwierig oder auch gar nicht durchführbar, letzteres insbesondere bei nur fossil bekannten Arten, die ja weder Kreuzungsexperimente noch Verhaltensbeobachtung zulassen. Aber auch Kreuzungsexperimente bringen nicht immer Sicherheit; abgesehen von dem Aufwand, den sie erfordern können, und ihrer sehr häufigen technischen Unmöglichkeit, ist es gut möglich, daß die zur Entscheidung erforderliche Generationenzahl in Gefangenschaft entweder nicht erreicht werden kann oder aber aufgrund der künstlich verbesserten Überlebensbedingungen gegenüber der Natur überschritten wird. (Trotzdem ermöglichen Zuchtexperimente sehr oft eindeutige Entscheidungen, z.B. dann, wenn die Nachkommen bestimmter Kreuzungen regelmäßig steril sind; das bekannteste Beispiel hierfür liefern der Allgemeinheit Pferd [Equus przewalskii] und Esel [E. africanus], bei Schmetterlingskundlern sind es das Große Wiener und das Atlantische Nachtpfauenauge [Saturnia pyri und S. atlantica].) Eine sichere Entscheidung erfordert daher in vielen Fällen langfristige, bei schwer erkennbaren Merkmalen am besten durch genetische Untersuchungen abgestützte Freilandbeobachtungen.
Arten mit einem großen Verbreitungsgebiet zerfallen oft infolge geographischer Barrieren in relativ geschlossene Fortpflanzungsgruppen, innerhalb derer sich der größte Teil der Paarungen vollzieht, während Paarungen zwischen Angehörigen verschiedener dieser Fortpflanzungsgruppen selten sind. Diese Gruppen heißen Populationen. Bilden sie stabile separate Merkmale aus, welche alle ihre Angehörigen aufweisen, diejenigen anderer Populationen aber nicht oder nur in einer abweichenden stabilen Kombination mit anderen stabilen Merkmalen, so heißen sie Unterarten oder Rassen. Jede Unterart hat also als Population angefangen, jede Art als Unterart, vorausgesetzt, wir erkennen jeden dauerhaften Mechanismus, der zur Paarungsverhinderung führt, neben dem geographischen (der auf jeden Fall der häufigste ist), als gleichwertig mit diesem an. Neben die geographischen Rassen können daher, wie oben ausgeführt, z.B. durch den Schlupfzeitpunkt getrennte (»phänologische« – von gr. 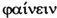 »erscheinen«) Rassen treten, die zu Arten werden können (z.B. bei Eupithecia innotata und E. unedonata, kleinen Schmetterlingen aus der Familie der Spanner. Verzögert man künstlich den Schlupf der früher erscheinenden Art, ist sie mit der späteren kreuzbar; aber die Nachkommen haben wieder getrennte Schlupfzeiten). Alle Populationen, deren Mitglieder mit den Mitgliedern anderer Populationen überhaupt keine Paarungen vollziehen, werden im Laufe unterschiedlich langer Zeit zu Arten. (Die Unterschiedlichkeit dieser Zeit wird durch mehrere Faktoren bestimmt, z.B.: die Unterschiedlichkeit der Umwelt [diese kann die Träger unterschiedlicher Eigenschaften hinsichtlich ihrer Überlebens- und daher Fortpflanzungswahrscheinlichkeit begünstigen]; die Generationslänge bzw. Lebenszeit; die Anzahl der Individuen; die strukturell, d.h. molekularanatomisch bedingt unterschiedliche Wahrscheinlichkeit von Veränderungen des Erbguts.) Wir können diese Unterschiedlichkeit in der Geschwindigkeit der Artbildung ausgezeichnet an den Eiszeitfolgen beobachten: diese Klimaveränderung ließ sehr viele Arten der Nordhalbkugel ihre Verbreitung nach Süden verlagern, wodurch aufgrund der geographischen Gegebenheiten mindestens zwei Populationen entstanden, zwischen deren Mitgliedern keine Paarungen mehr möglich waren. Mit dem Rückgang der Gletscher wanderten diese zuvor getrennten Populationen oft wieder nordwärts, wodurch Paarungen zwischen ihren verschiedenen Mitgliedern wieder möglich wurden. Auch wenn die getrennten Populationen inzwischen zu Unterarten geworden waren, so hoben sich ihre inzwischen erworbenen Unterschiede allmählich wieder auf (wir können diesen Vorgang bei einigen Laufkäferarten der Gattung Carabus noch nachweisen; bei den meisten anderen betroffenen Arten dürfte der Nachweis nicht mehr möglich sein). Sie konnten aber auch zu Arten geworden sein, und dann blieben diese notwendigerweise konstant. (Das bekannteste Beispiel dafür bilden die Rotbauch- und die Gelbbauchunke.) Sie können aber auch schneller Eigenschaften erwerben, die ihre Überlebenswahrscheinlichkeit in dem sich erwärmenden südlichen Gebiet erhöhen, das sie inzwischen bewohnen, als sich die Gletscher zurückziehen, und daher in ihrem neuen, zerrissenen Verbreitungsgebiet bleiben und dieses nicht nach Norden zurückverlegen; bekanntestes Beispiel hierfür ist die Blauelster (Cyanopica cyanus), welche ausschließlich in Südchina und Südportugal vorkommt. (Da von ihr in Portugal über 10.000 Jahre alte Fossilien gefunden wurden, scheidet eine Verschleppung nach Portugal, z.B. via Macao, als Ursache dieses Verbreitungsmusters aus.) Beide Populationen sind nach Aussehen (»morphologisch«) und Erbgut (»genetisch«) zwar nachweislich, aber nur extrem geringfügig unterschieden; ihre Vereinigung ergäbe wahrscheinlich sehr schnell die Aufhebung aller Unterschiede.
»erscheinen«) Rassen treten, die zu Arten werden können (z.B. bei Eupithecia innotata und E. unedonata, kleinen Schmetterlingen aus der Familie der Spanner. Verzögert man künstlich den Schlupf der früher erscheinenden Art, ist sie mit der späteren kreuzbar; aber die Nachkommen haben wieder getrennte Schlupfzeiten). Alle Populationen, deren Mitglieder mit den Mitgliedern anderer Populationen überhaupt keine Paarungen vollziehen, werden im Laufe unterschiedlich langer Zeit zu Arten. (Die Unterschiedlichkeit dieser Zeit wird durch mehrere Faktoren bestimmt, z.B.: die Unterschiedlichkeit der Umwelt [diese kann die Träger unterschiedlicher Eigenschaften hinsichtlich ihrer Überlebens- und daher Fortpflanzungswahrscheinlichkeit begünstigen]; die Generationslänge bzw. Lebenszeit; die Anzahl der Individuen; die strukturell, d.h. molekularanatomisch bedingt unterschiedliche Wahrscheinlichkeit von Veränderungen des Erbguts.) Wir können diese Unterschiedlichkeit in der Geschwindigkeit der Artbildung ausgezeichnet an den Eiszeitfolgen beobachten: diese Klimaveränderung ließ sehr viele Arten der Nordhalbkugel ihre Verbreitung nach Süden verlagern, wodurch aufgrund der geographischen Gegebenheiten mindestens zwei Populationen entstanden, zwischen deren Mitgliedern keine Paarungen mehr möglich waren. Mit dem Rückgang der Gletscher wanderten diese zuvor getrennten Populationen oft wieder nordwärts, wodurch Paarungen zwischen ihren verschiedenen Mitgliedern wieder möglich wurden. Auch wenn die getrennten Populationen inzwischen zu Unterarten geworden waren, so hoben sich ihre inzwischen erworbenen Unterschiede allmählich wieder auf (wir können diesen Vorgang bei einigen Laufkäferarten der Gattung Carabus noch nachweisen; bei den meisten anderen betroffenen Arten dürfte der Nachweis nicht mehr möglich sein). Sie konnten aber auch zu Arten geworden sein, und dann blieben diese notwendigerweise konstant. (Das bekannteste Beispiel dafür bilden die Rotbauch- und die Gelbbauchunke.) Sie können aber auch schneller Eigenschaften erwerben, die ihre Überlebenswahrscheinlichkeit in dem sich erwärmenden südlichen Gebiet erhöhen, das sie inzwischen bewohnen, als sich die Gletscher zurückziehen, und daher in ihrem neuen, zerrissenen Verbreitungsgebiet bleiben und dieses nicht nach Norden zurückverlegen; bekanntestes Beispiel hierfür ist die Blauelster (Cyanopica cyanus), welche ausschließlich in Südchina und Südportugal vorkommt. (Da von ihr in Portugal über 10.000 Jahre alte Fossilien gefunden wurden, scheidet eine Verschleppung nach Portugal, z.B. via Macao, als Ursache dieses Verbreitungsmusters aus.) Beide Populationen sind nach Aussehen (»morphologisch«) und Erbgut (»genetisch«) zwar nachweislich, aber nur extrem geringfügig unterschieden; ihre Vereinigung ergäbe wahrscheinlich sehr schnell die Aufhebung aller Unterschiede.
Arten, die ein sehr großes Verbreitungsgebiet besitzen, können in mehrere Unterarten zerfallen, die mit ihren näheren Nachbarn problemlos kreuzbar sind (Rassenkreis), nicht aber mit ihren entferntesten. Diese können, wenn der Rassenkreis z.B. den Erdball umrundet, durchaus am gleichen Ort wie die ursprünglich entfernteste Rasse auftreten (sympatrisch auftreten); in diesem Fall verhalten sie sich ihr gegenüber wie eine andere Art. (Bekanntestes Beispiel hierfür sind die Silbermöwe und die Heringsmöwe, welche die jeweiligen Endpunkte eines Rassenkreises bilden.) Man spricht in diesem Fall von Halbarten oder Semispezies. (Zwei normale getrennte Arten können also auch durch Aussterben verbindender Unterarten eines Rassenkreises entstehen.) Die Art ist die fundamentale Einheit des Natürlichen Systems der Lebewesen; dessen Erforschung widmet sich die Spezielle Biologie (Spezielle Zoologie, Spezielle Botanik u.ä.), denn der internationale lateinische Ausdruck für »Art« lautet species. Die species ist daher das fundamentale Taxon des Natürlichen Systems. Mehrere (oft nur mutmaßlich) auf die gleiche Ausgangsart zurückführbare (verwandte) Arten werden in einer Gattung zusammengefaßt; die Gattung (genus) ist also gegenüber der Art (species) das höhere Taxon. Verwandte Gattungen lassen sich zu einer Familie (familia) zusammenfassen – einem noch höheren Taxon als die Gattung also –, verwandte Familien zu einer Ordnung (ordo), verwandte Ordnungen zu einer Klasse (classis), verwandte Klassen zu einem Stamm (phylum), verwandte Stämme zu einem Reich (regnum) usw. Diese Taxa lassen sich sowohl aufgliedern wie zusammenfassen, z.B. zu Unterfamilien, Überfamilien, Unterklassen, Überreichen u.v.a.
Da, wie wir schon gelernt haben, jede Art (den Klonen genügt für ein neues Art-Analogon ein einzelnes Individuum) nur aus einer anderen Art durch Trennung der Fortpflanzungsgemeinschaft hervorgehen kann, sollte jedes höhere Taxon erstens nur die Arten erfassen, die von einer Stammart abstammen (monophyletisch sein), zweitens alle diese (beliebig vielen) Arten vollständig. (Da das Leben auf der Erde höchstwahrscheinlich nur einmal entstanden ist, stammen also alle Arten – wenn wir den Begriff auf die Klone ausweiten – sämtlich von einer einzigen Art ab!) Oft wird fälschlich behauptet, keine lebende Art sei die Stammart einer anderen lebenden Art; in Wirklichkeit können sich von einer z.B. durch die große Zahl ihrer Mitglieder unveränderten Art sehr wohl sogar mehrere kleine Populationen, etwa auf Inseln, abgespalten und zu Arten entwickelt haben. Dies ist beispielsweise der Fall bei der Ringeltaube (Columba palumbus), einer weitverbreiteten Art, von welcher sich nicht weniger als drei weitere lebende Arten abgespalten haben, die makaronesischen Lorbeertauben (C. trocaz, C. bollii u. C. iunoniae), von welchen zwei sogar, untrüglichstes Zeichen abgeschlossener Artbildung, am gleichen Ort (sympatrisch) vorkommen (und inzwischen sogar, sekundär nämlich, durch menschliches Zutun, sympatrisch mit der Art, von der sie abstammen, d.h. der Ringeltaube). Nahe verwandte sympatrische Arten haben übrigens immer – denn sonst verdrängt die eine die andere, läßt sie also aussterben – eine etwas unterschiedliche Lebensweise, besetzen also verschiedene ökologische Nischen. (Allerdings gilt dies für alle an einem Ort koexistierenden Arten; nur läßt es sich bei nahe verwandten besonders augenfällig beobachten.)
Da jede Art nur durch Trennung von einer schon bestehenden Art entstehen kann, nämlich durch Trennung von der Fortpflanzungsgemeinschaft, beginnt jedes neue höhere Taxon mit einer von einer anderen abgespaltenen Art (welche dann Schwesterart, engl. sibling species, der anderen genannt wird); sowohl aus dieser wie aus jener können im Laufe der Zeit beliebig viele neue Arten entstehen, deren Gesamtheit im Natürlichen System dann das Schwestertaxon der anderen Gesamtheit bildet. Beide höheren Taxa müssen dann also, ungeachtet ihrer Größe (sie können auch nur eine einzige Art enthalten; dann heißen sie monotypisch), die gleiche Ranghöhe zugeteilt bekommen, also beide jeweils eine Familie, Unterordnung, Überklasse usw. sein; in jedem anderen Fall ist ihre Einordnung falsch. Es gibt also nur eine richtige Abbildung des Natürlichen Systems; ihrer Ermittlung, die selbstverständlich aufgrund Datenmangels für alle Zeiten nur annähernd geleistet werden kann, widmen sich die Systematiker. Das Natürliche System zeichnet sich also durch das aus, was der englische Biologe Dawkins die »perfekte Einschachtelung« nennt: diese ist also nicht etwa durch die ästhetischen oder praktischen (z.B. Übersichts-) Bedürfnisse der Speziellen Biologen bewirkt worden, sondern ist die unvermeidliche Folge der – durch sie nur möglichst genau abgebildeten – Abstammungsverhältnisse.
Das Natürliche System mit seiner Benennungsordnung wurde verbindlich 1758 von dem schwedischen Botaniker Carl von Linné entwickelt; von ihm stammt sowohl der Grundstock der taxonomischen Kategorien wie auch die Regeln der Artbezeichnung (Nomenklatur; dort Einzelheiten): jede Art erhält einen kleingeschriebenen Namen in Latein oder latinisiertem Griechisch (oder einen, der einigermaßen als solches gelten kann), welcher hinter den großgeschriebenen, ebenfalls auf die genannte Art gebildeten Gattungsnamen gesetzt wird (binäre Nomenklatur). Jeder Artname darf nur einmal in einer Gattung, jeder Gattungsname nur einmal im ganzen Natürlichen System vorkommen (faktisch duldet man allerdings den gleichen Namen für je eine Tier- und eine Pflanzengattung; dagegen hat sich, besonders aus den USA, Opposition erhoben). Im Zusammenhang mit der obligatorisch mit dem Namen verbundenen, bestimmten Regeln unterworfenen Beschreibung ist dadurch jede Art eindeutig identifizierbar (und nationale Rivalitäten ausgeschaltet). Beispielsweise heißt der Braunbär »Ursus arctos«, wobei »Ursus«, der die Gattung bezeichnet, das lateinische Wort für »Bär« ist, »arctos« das latinisierte (also der lateinischen Orthographie angepaßte) griechische Wort für »Bär«. Der Eisbär, welcher der gleichen Gattung angehört, heißt Ursus maritimus (also »Meerbär«). Nun hat der Braunbär Rassen gebildet, z.B. den Grizzly; zur Bezeichnung der Rasse wird ein weiterer Bestandteil an den Namen angehängt, in unserem Fall »horribilis« (will man sichergehen, daß diese Benennung eine Rasse und nicht etwa eine Morphe oder eine ökologische Variante bezeichnet, so schiebt man vor dem dritten Namenselement ein »ssp.« = subspecies ein [also: Ursus arctos <ssp.> horribilis, »der schreckliche Braunbär« bzw. »der schreckliche Bär Bär«]; heute ist dieser Einschub nicht mehr gebräuchlich, da die Internationale Nomenklaturkommission die Morphen- und Variantenbezeichnungen nicht mehr schützt). Aus der binären Nomenklatur ist also für (nicht durch menschliche Zucht entstandene) Rassen = Unterarten eine ternäre Nomenklatur geworden. Noch weitergehende Aufspaltungen, die sich in fester Namensgebung niederschlagen, sind in der Speziellen Biologie nicht mehr gebräuchlich; die Fortschritte der Genomanalyse haben sie zu oft unhandlich bis überflüssig werden lassen.
Übersichtsbildende Literatur: Richard Dawkins, Das egoistische Gen, Berlin/HD/N.Y. 1978; Ernst Mayr, Eine neue Philosophie der Biologie, München/Zürich 1991. Weiterführend: J.W.Wägele, Grundlagen der Phylogenetischen Systematik, München (Pfeil) 2000




 zurück
zurück

